У кожній еукаріотичній клітині ховається складна система енергетичних і регуляторних структур, оточених не однією, а двома мембранами. Ці органели створюють ізольовані простори, де хімічні процеси відбуваються з максимальною ефективністю — від виробництва універсальної енергетичної молекули АТФ до синтезу органічних сполук під впливом світла. Мітохондрії та хлоропласти (як основні представники пластид) — це не просто «станції» чи «фабрики». Вони є результатом давніх еволюційних подій, що назавжди змінили хід життя на Землі.
Для тих, хто тільки знайомиться з темою: двомембранні органели — це мітохондрії, які забезпечують клітинне дихання й постачання енергії майже в усіх еукаріотах, та хлоропласти в рослинних клітинах, відповідальні за фотосинтез. Просунуті читачі знайдуть тут детальний розбір мембранних комплексів, транспортних механізмів, доказів ендосимбіотичного походження та зв’язків із реальними захворюваннями й біотехнологіями.
Основні двомембранні органели та їхнє місце в клітині
Мітохондрії присутні майже в усіх еукаріотичних клітинах — від найпростіших до клітин людини. Хлоропласти характерні для рослин і деяких водоростей. Обидві органели мають зовнішню та внутрішню мембрани, міжмембранний простір і внутрішній матрикс (або строму). Подвійна мембрана дозволяє підтримувати різні середовища та створювати електрохімічні градієнти, без яких ефективне виробництво енергії було б неможливим.
Ядерна оболонка також складається з двох мембран, але традиційно в українській біологічній освіті до класичних двомембранних органел відносять саме мітохондрії та пластиди. Ядерна оболонка виконує захисну й регуляторну роль, відокремлюючи генетичний матеріал, тоді як мітохондрії та хлоропласти безпосередньо пов’язані з енергетичним обміном.
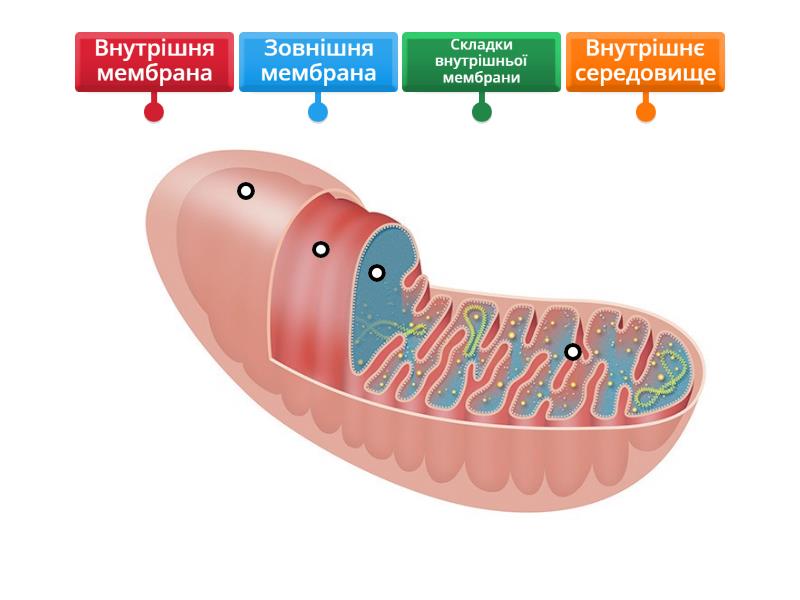
Мітохондрії: мікроскопічні електростанції з гнучкою архітектурою
Мітохондрія має довгасту або овальну форму завдовжки 2–10 мікрометрів. Зовнішня мембрана гладка й пронизана поринами — білковими каналами, що пропускають молекули до 5 кДа. Вона нагадує сито, яке захищає внутрішній вміст, але дозволяє вільний обмін невеликих речовин з цитоплазмою.
Внутрішня мембрана непроникна для більшості іонів і молекул. Саме вона утворює численні вирости — кристи. У різних клітинах форма крист відрізняється: у клітинах печінки вони часто пластинчасті, у м’язових — трубчасті або ламелярні. Кристи різко збільшують площу поверхні, де розміщуються комплекси електрон-транспортного ланцюга (I–IV) та АТФ-синтаза. На внутрішній мембрані також локалізований унікальний ліпід — кардіоліпін, який стабілізує суперкомплекси дихального ланцюга.
Між зовнішньою та внутрішньою мембранами лежить міжмембранний простір — невеликий, але критично важливий резервуар протонів. У матриксі (внутрішньому просторі) відбуваються реакції циклу Кребса, тут містяться мітохондріальна ДНК (кільцева, як у бактерій), рибосоми 70S типу та ферменти. Мітохондріальна ДНК людини кодує лише 13 білків, решту — ядерні гени, що підкреслює глибоку інтеграцію органели з клітиною.
Процес окислювального фосфорилювання розгортається поетапно. Піруват або жирні кислоти надходять у матрикс, де окислюються. Електрони рухаються ланцюгом комплексів, «викачуючи» протони в міжмембранний простір. Коли протони повертаються через АТФ-синтазу, енергія градієнта перетворюється на АТФ. Одна молекула глюкози в аеробних умовах дає до 30–32 молекул АТФ — у десятки разів більше, ніж гліколіз у цитоплазмі.
Мітохондрії динамічні: вони постійно зливаються (фузія) та діляться (фіссія). Цей цикл дозволяє видаляти пошкоджені ділянки через мітопігію та адаптувати кількість органел до енергетичних потреб клітини. У нейронах мітохондрії переміщуються аксоном на відстань до метра, живлячи синапси. У сперматозоїдах їх небагато, але вони критичні для рухливості. У яйцеклітині людини їх кількість сягає сотень тисяч — запас на ранні етапи ембріогенезу.
Порушення роботи мітохондрій лежать в основі цілої групи захворювань. Мутації в мітохондріальній ДНК або ядерних генах, що кодують мітохондріальні білки, викликають синдроми Лея, MELAS, MERRF, спадкову оптичну нейропатію Лебера. Симптоми часто зачіпають тканини з високим енергоспоживанням: м’язи, мозок, серце, зір. Деякі препарати (наприклад, певні антибіотики) можуть тимчасово пригнічувати мітохондріальні рибосоми через їхню схожість із бактеріальними.
Хлоропласти та інші пластиди: сонячні лабораторії рослинного світу
Хлоропласт — це дископодібна органела завбільшки 5–10 мікрометрів, оточена двома мембранами. Зовнішня мембрана гладка, внутрішня утворює систему впинань. У стромі (аналог матриксу) відбуваються реакції темнової фази фотосинтезу — цикл Кальвіна. Тут містяться кільцева ДНК хлоропласту, рибосоми 70S та ферменти.
Світлова фаза розгортається на тилакоїдних мембранах. Тилакоїди — плоскі мішечки, які в місцях щільного укладання утворюють грани (стопки, що нагадують монети). Між гранами тилакоїди з’єднані стромальними ламелами. На мембранах тилакоїдів розташовані фотосистеми I та II, цитохромний комплекс і АТФ-синтаза. Хлорофіл та допоміжні пігменти вловлюють фотони, запускаючи потік електронів. У результаті утворюються АТФ, НАДФН і кисень як побічний продукт.
Хлоропласти — лише один тип пластид. З пропластидів можуть розвиватися хромопласти (накопичують каротиноїди, забарвлюють плоди та пелюстки), амілопласти (запасають крохмаль у коренях і бульбах), лейкопласти (безбарвні, синтезують жирні кислоти). У деяких водоростей вторинний ендосимбіоз призвів до появи хлоропластів з трьома або чотирма мембранами.
Фотосинтез не просто «виробляє кисень». Він фіксує вуглець, формує основу харчових ланцюгів і регулює атмосферний склад. У листкових клітинах мезофілу хлоропласти можуть займати до половини об’єму цитоплазми. Вони здатні переміщуватися вздовж цитоскелету, оптимізуючи положення відносно світла (фототаксис).
Порівняння мітохондрій та хлоропластів
| Характеристика | Мітохондрії | Хлоропласти |
|---|---|---|
| Кількість мембран | Дві | Дві (у вищих рослин) |
| Внутрішні мембранні структури | Кристи | Тилакоїди (грани + стромальні ламели) |
| Основний процес | Окислювальне фосфорилювання (АТФ) | Фотосинтез (АТФ + НАДФН + органічний вуглець) |
| Власний геном | Кільцева ДНК (~16,5 тис. пар нуклеотидів у людини) | Кільцева ДНК (120–160 тис. пар нуклеотидів) |
| Рибосоми | 70S | 70S |
| Походження (ендосимбіотична теорія) | Альфа-протеобактерії | Ціанобактерії |
| Присутність у клітинах | Майже всі еукаріоти | Рослини, водорості, деякі протести |
Ендосимбіотична теорія: як два прокаріотичні світи подарували еукаріотам енергетичну незалежність
Найпереконливіше пояснення походження мітохондрій та хлоропластів дає ендосимбіотична теорія. Приблизно 1,5–2 мільярди років тому архейна клітина-господар поглинула аеробну альфа-протеобактерію. Замість того щоб перетравити її, господар зберіг симбіонта. Бактерія отримала захист і стабільне середовище, а господар — ефективний механізм аеробного дихання. Пізніше, вже в лінії рослин, подібна подія сталася з ціанобактерією — так з’явилися хлоропласти.
Докази вражають своєю багатогранністю. Обидві органели мають власну кільцеву ДНК, рибосоми бактеріального типу (70S), діляться бінарним поділом незалежно від клітинного циклу. Внутрішня мембрана за складом ліпідів і білків ближча до бактеріальної плазматичної мембрани. Філогенетичний аналіз показує, що мітохондріальні гени closest до генів Rickettsia та інших альфа-протеобактерій, а хлоропластні — до ціанобактерій. Масштабний перенос генів з органел у ядро зробив еукаріотичну клітину цілісною системою, але зберіг «пам’ять» про незалежне минуле.
У деяких сучасних організмів процеси ендосимбіозу тривають. Наприклад, у певних інфузорій або молюсків спостерігаються «вторинні» або навіть «третинні» хлоропласти. Це показує, що еволюція не зупинилася — вона продовжує експериментувати з симбіотичними партнерствами.
Ендосимбіотична теорія пояснює не лише будову двох органел — вона розкриває фундаментальний механізм, завдяки якому прості прокаріотичні клітини змогли створити складні багатоклітинні організми з високим енергетичним бюджетом.
Цікаві факти про двомембранні органели
Цікаві факти
- У середній клітині людини налічується від кількох сотень до кількох тисяч мітохондрій. У зрілій яйцеклітині їх кількість може сягати сотень тисяч — це стратегічний енергетичний запас для перших поділів ембріона.
- Мітохондріальна ДНК передається майже виключно по материнській лінії. Сперматозоїд вносить у яйцеклітину лише кілька мітохондрій, які зазвичай деградують після запліднення.
- Кристи мітохондрій у клітинах з високим енергоспоживанням (серцевий м’яз, скелетні м’язи) можуть займати до 40 % внутрішнього об’єму органели.
- Хлоропласти здатні «переїжджати» всередині клітини вздовж актинових філаментів, оптимізуючи освітлення. У тіні вони скупчуються біля верхньої стінки, на яскравому світлі — розподіляються рівномірніше.
- Деякі антибіотики (хлорамфенікол, тетрацикліни) пригнічують синтез білка в мітохондріях і хлоропластах, бо їхні рибосоми схожі на бактеріальні. Це пояснює певні побічні ефекти терапії.
- У червоних водоростей і багатьох протистів хлоропласти оточені трьома або чотирма мембранами — наслідок вторинного ендосимбіозу, коли еукаріотична клітина поглинула іншу еукаріотичну клітину з уже наявним хлоропластом.
Практичне значення та сучасні горизонти
Розуміння двомембранних органел безпосередньо впливає на медицину та сільське господарство. Мітохондріальна дисфункція пов’язана не лише з рідкісними генетичними синдромами, а й із процесами старіння, нейродегенеративними захворюваннями, цукровим діабетом 2 типу та онкологією. Дослідження біогенезу мітохондрій (через фактори PGC-1α) відкривають шляхи до терапевтичного впливу на м’язову витривалість і метаболізм.
У рослинництві інженерія хлоропластів дозволяє підвищувати ефективність фотосинтезу, стійкість до посухи чи створювати рослини з покращеним поживним складом. Експерименти з перенесенням генів між органелами та ядром тривають. Мітохондріальна замісна терапія (технологія «дитини від трьох батьків») вже допомогла родинам уникнути передачі тяжких мітохондріальних захворювань.
Кожна нова деталь будови крист чи тилакоїдів, кожен уточнений механізм транспорту протонів чи електронів — це крок до більш точного втручання в енергетичний баланс клітини. Двомембранні органели залишаються одними з найелегантніших прикладів того, як еволюція перетворює випадкові симбіотичні події на стійкі, високоефективні біологічні системи, що підтримують життя в усіх його проявах.